
De especies silvestres a razas locales y cultivos

Patrick F. Byrne, Dept. of Soil and Crop Sciences, Colorado State University, Fort Collins, Colorado 80523 (Patrick.Byrne@colostate.edu)
Christopher Richards, USDA-ARS National Laboratory for Genetic Resources Preservation, 1111 S. Mason St., Fort Collins, Colorado, 80521
Gayle M. Volk, USDA-ARS National Laboratory for Genetic Resources Preservation, 1111 S. Mason St., Fort Collins, Colorado, 80521
El propósito de este capítulo es proporcionar una descripción general del proceso de domesticación desde especies silvestres hasta la domesticación temprana (razas locales) y su posterior mejora a los cultivos modernos.
Contenido
- Introducción
- Domesticación temprana de cultivos
- Breve historia del fitomejoramiento
- Referencias
- Agradecimientos
1. Introducción
Generalmente, los parientes silvestres de las especies de plantas domesticadas no poseen todos los rasgos deseables para la producción y el uso agrícola de rutina. Este capítulo ofrece una descripción general del proceso de domesticación temprana de cultivos desde hace miles de años hasta el progreso en el mejoramiento de plantas por parte de los fitomejoradores de hoy en día. La comprensión de la domesticación de cultivos y el fitomejoramiento proporciona un contexto para el valor, la importancia y el uso de parientes silvestres de cultivos que se conservan in situ o en bancos de germoplasma.
2. Domesticación temprana de cultivos
Uno de los hitos de la historia humana es la transición de sociedades nómadas de cazadores-recolectores a sociedades asentadas basadas en la agricultura, la llamada revolución neolítica. Un componente clave de esta transición fue la domesticación de especies silvestres en cultivos capaces de soportar densidades de población más altas (Harlan, 1992; Schwanitz, 1966). La domesticación de cultivos a partir de especies silvestres comenzó hace unos 8,000 a 10,000 años de forma independiente en muchas partes del mundo. Algunos de los centros de domesticación más notables fueron el Creciente Fértil del Medio Oriente (trigo, cebada, lenteja, y garbanzo), Mesoamérica (maíz, chiles, calabaza, y frijol común), la región Andina (papa, tomate y un segundo centro de origen para el frijol común) y el Sudeste asiático (arroz, mijo, y soya).
Ya sea de forma deliberada o no, los primeros agricultores seleccionaron con base en rasgos que hicieron que las plantas silvestres fueran más adecuadas para las necesidades humanas (Figura 1). Estos incluyeron rasgos que mejoraron el rendimiento (mayor tamaño del producto cosechado; menor ramificación), facilitaron la producción agrícola (pérdida de latencia de la semilla; retención de la semilla en la planta) o mejoraron la calidad del producto (reducción del amargor o propiedades tóxicas; menor dificultad de procesamiento). El término “síndrome de domesticación” se ha utilizado para abarcar la colección de rasgos que se seleccionaron de forma común (Gepts, 2018).
 Figura 1. Comparación de semillas de especies progenitoras silvestres (izquierda) y sus contrapartes domesticadas (derecha). Se observa el cambio en el tamaño de la semilla, así como la reducción de la pigmentación en algunos casos. De arriba a abajo: pistacho, café, soya, cebada, arroz silvestre norteamericano (Zizania palustris) y sorgo. Crédito de la foto: Christina Walters.
Figura 1. Comparación de semillas de especies progenitoras silvestres (izquierda) y sus contrapartes domesticadas (derecha). Se observa el cambio en el tamaño de la semilla, así como la reducción de la pigmentación en algunos casos. De arriba a abajo: pistacho, café, soya, cebada, arroz silvestre norteamericano (Zizania palustris) y sorgo. Crédito de la foto: Christina Walters.
Muchos de los rasgos de domesticación se caracterizan por tener herencia simple, es decir, están controlados por uno o dos genes con grandes efectos en lugar de múltiples genes. La herencia simple de rasgos de domesticación facilitó su selección e incorporación a poblaciones silvestres (Gepts, 2018). Lo más probable es que la domesticación haya ocurrido durante un período prolongado, con un proceso continuo de polinización cruzada entre poblaciones silvestres y domesticadas. Como las plantas seleccionadas representaban solo una pequeña fracción de la población silvestre total, a menudo la domesticación estuvo acompañada de una reducción de la diversidad genética (ver Figura 3). Algunos de los rasgos involucrados en la transición del teosinte silvestre al maíz domesticado se muestran en el Video 1, “Domesticación del maíz”.
Video 1. Domesticación del maíz. (Subtítulos disponibles en español).
A medida que los primeros agricultores emigraron a nuevos territorios, trajeron consigo sus semillas, tubérculos y portainjertos. Los cultivos encontraron una variedad de tipos de suelo, patrones climáticos, plagas de insectos, y patógenos. En respuesta a los nuevos ambientes, los agricultores seleccionaron plantas que se adaptaron mejor a las condiciones locales. Así surgieron las “variedades locales”, variedades tradicionales seleccionadas por los agricultores para adaptarse a las condiciones locales y a las preferencias alimentarias. Por lo general, estas variedades tienen bajo potencial de rendimiento, pero a menudo son genéticamente diversas y resistentes al estrés. Muchas de las denominadas ‘variedades vestigiales’ son ejemplos de variedades locales. Normalmente, las variedades locales son genéticamente heterogéneas (no uniformes), y por lo tanto, son una fuente importante de diversidad genética para el mejoramiento de cultivos. Las variedades locales suelen ser menos diversas que sus ancestros silvestres, pero más diversas que los cultivos producidos durante la era moderna del fitomejoramiento. Esto se muestra conceptualmente en la Figura 2 y en la base del análisis de ADN en frijol común (Figura 3).
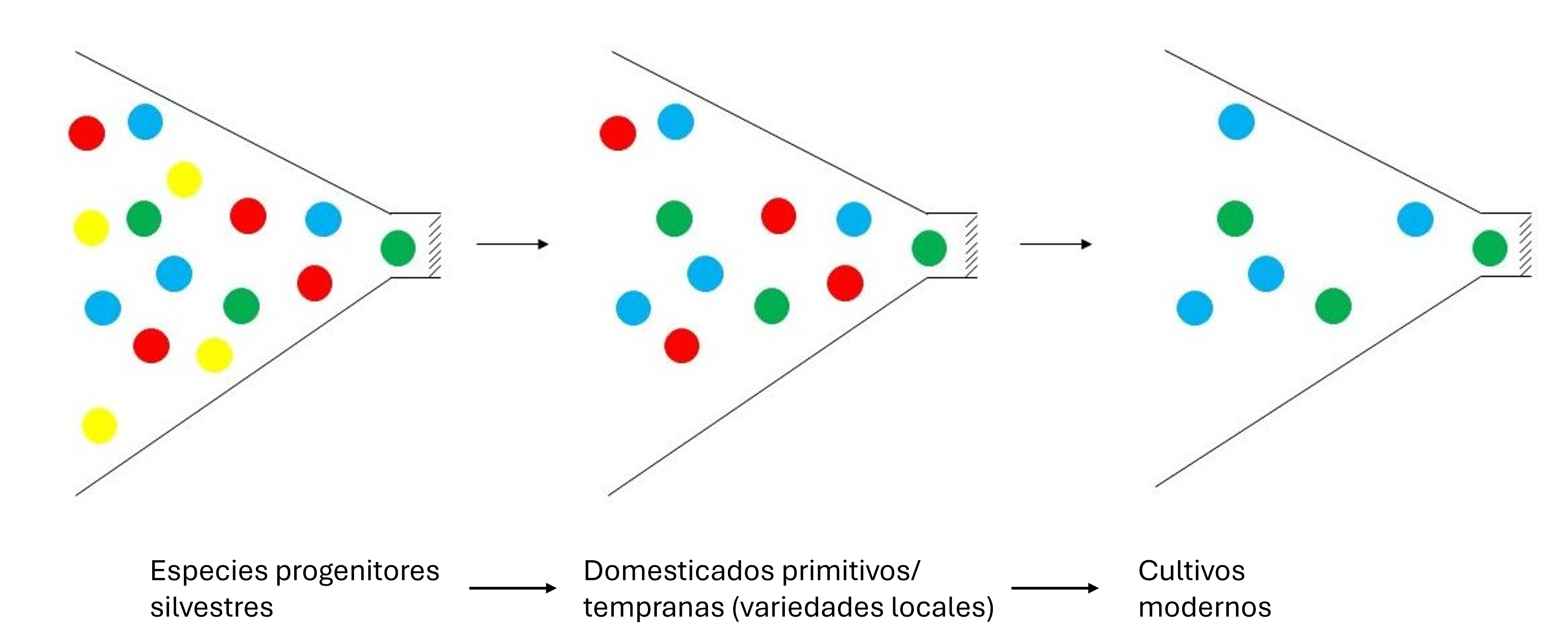 Figura 2. Representación conceptual de la reducción en la diversidad genética (efecto de cuello de botella) a medida que las especies silvestres fueron domesticadas y de la reducción adicional ocurrida con el desarrollo de cultivos modernos. Los colores representan la diversidad de alelos dentro de una población. Rediseñado de Tanksley y McCouch (1997). En lugar de cambios abruptos como lo indican aquí los pronunciados cuellos de botella, ahora se considera que la domesticación y el mejoramiento de cultivos conducen a una pérdida gradual de la diversidad durante un largo período de tiempo.
Figura 2. Representación conceptual de la reducción en la diversidad genética (efecto de cuello de botella) a medida que las especies silvestres fueron domesticadas y de la reducción adicional ocurrida con el desarrollo de cultivos modernos. Los colores representan la diversidad de alelos dentro de una población. Rediseñado de Tanksley y McCouch (1997). En lugar de cambios abruptos como lo indican aquí los pronunciados cuellos de botella, ahora se considera que la domesticación y el mejoramiento de cultivos conducen a una pérdida gradual de la diversidad durante un largo período de tiempo.
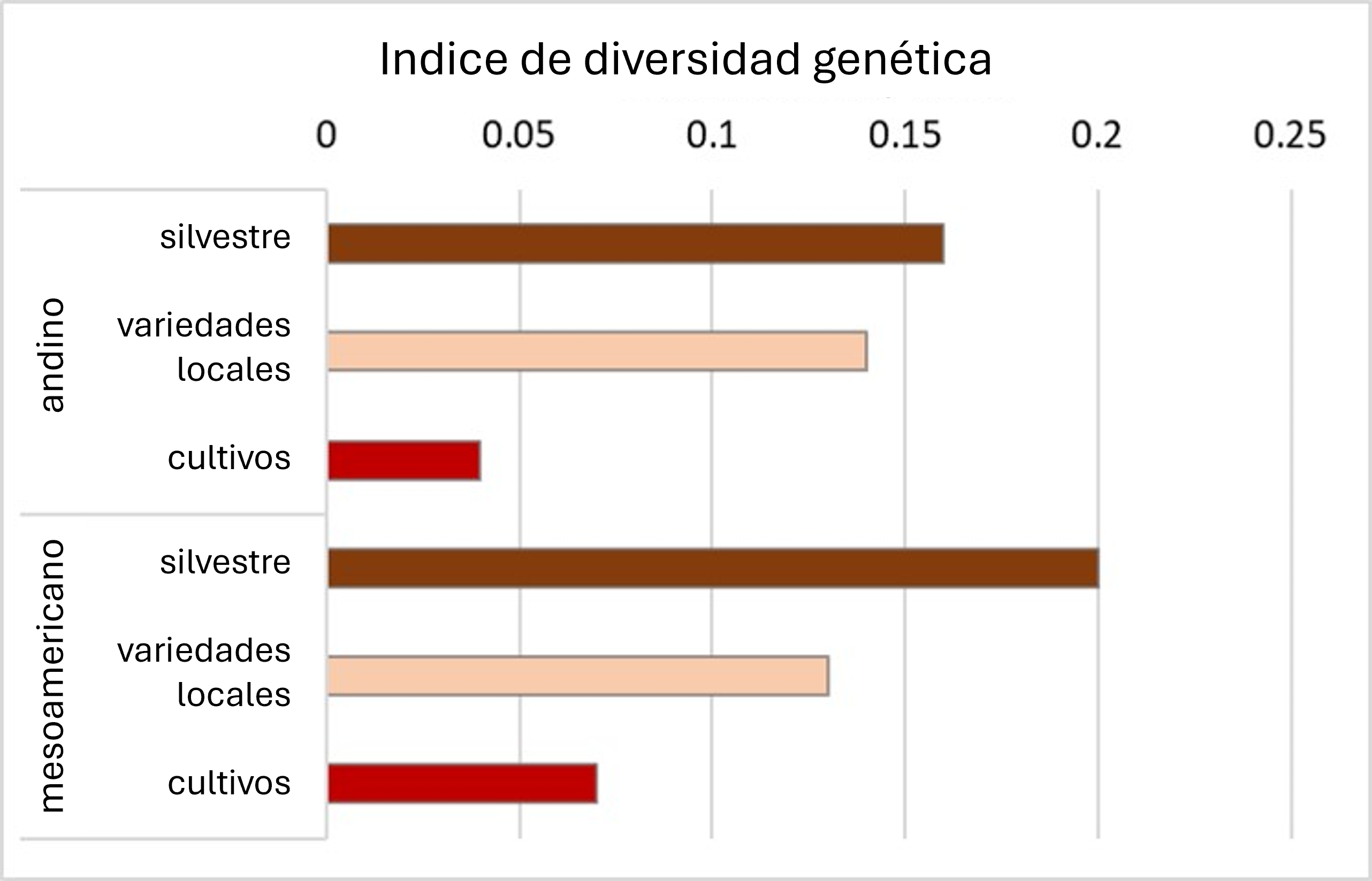 Figura 3. Reducción de la diversidad genética durante la evolución del frijol común (Phaseolus vulgaris) en los centros de origen andino y mesoamericano. Rediseñado de Gepts (2018).
Figura 3. Reducción de la diversidad genética durante la evolución del frijol común (Phaseolus vulgaris) en los centros de origen andino y mesoamericano. Rediseñado de Gepts (2018).
Video 2. ¿Cómo afecta la domesticación y el fitomejoramiento a la diversidad genética de las especies? (Subtítulos disponibles en español).
3. Breve historia del fitomejoramiento
El fitomejoramiento se ha definido como el “arte y la ciencia de mejorar la herencia de las plantas para beneficio de la humanidad” (Sleper y Poehlman, 2006). Los fitomejoradores participan en un proceso cíclico de recolección o generación de variación hereditaria de los rasgos de interés, evaluando dicha variación y seleccionando individuos superiores. El fitomejoramiento empírico o intuitivo fue practicado durante muchos siglos por agricultores observadores que reconocieron el valor de rasgos particulares y seleccionaron una parte de su cosecha para plantar la siguiente temporada (Murphy, 2007). Aunque no entendieron la base científica de la herencia de rasgos, lograron la mejora de los cultivos a través de sus selecciones.
El fitomejoramiento científico se remonta a alrededor de 1900, cuando se redescubrió el trabajo de Gregorio Mendel sobre la herencia de rasgos. Los fitomejoradores aplicaron los principios de Mendel a la selección de rasgos cualitativos (controlados por uno o dos genes principales) y desarrollaron métodos cada vez más sofisticados para mejorar los rasgos cuantitativos (controlados por muchos genes, cada uno con un efecto pequeño). El concepto de heredabilidad (la proporción de la variabilidad total debida a la genética en oposición a factores ambientales) se dilucidó y ayudó a orientar las estrategias de evaluación y selección. El diseño de parcelas de campo y los métodos de análisis estadístico mejoraron aún más la precisión con la que se evaluaron los rasgos y se compararon las variedades candidatas. La observación del vigor híbrido y su opuesta, la depresión endogámica, llevaron al desarrollo de la industria de semillas híbridas, inicialmente en maíz (Video 3) y luego en otros cultivos.
Video 3. Vigor híbrido en maíz. (Subtítulos disponibles en español).
A inicios de la década de 1960, la Revolución Verde transformó la producción de trigo y arroz mediante la incorporación de genes semi-enanizantes en variedades más altas. La altura reducida de la planta significaba que más fotosintatos se podían dirigir a la producción de granos en lugar de a tallos y hojas. Los tallos más cortos y rígidos también redujeron la cantidad de acame (doblamiento de los tallos). Aunque la Revolución Verde evitó la hambruna y salvó muchas vidas, no está exenta de controversia, como se describe en el informe del IFPRI (2002).
Otra revolución en el fitomejoramiento ocurrió con el desarrollo y aprovechamiento de marcadores moleculares de ADN a partir de la década de 1980 (Jain y Brar, 2010). Las tecnologías de marcadores moleculares permitieron: determinar relaciones genéticas (por ejemplo, entre cultivos y sus progenitores silvestres); obtener la huella genética de cultivos y líneas de mejoramiento; dilucidar la base bioquímica de la expresión de rasgos; localizar y caracterizar los loci de rasgos cuantitativos (QTL, por sus siglas en inglés, son genes con efectos pequeños que contribuyen a la herencia de rasgos cuantitativos) y realizar selección asistida por marcadores moleculares basada en patrones de ADN, en lugar de observación de rasgos. Los marcadores individuales son útiles cuando están muy relacionados con un gen principal. En la actualidad, muchos programas de mejoramiento aplican la selección genómica, por lo cual los efectos aditivos de miles de marcadores moleculares se incorporan en modelos de selección (Voss-Fels et al., 2019).
Aunque las tecnologías genómicas han permitido el alto rendimiento del genotipado de los patrones de ADN de las plantas, el fenotipado (evaluación de rasgos) de un gran número de líneas de mejoramiento sigue siendo un cuello de botella. Se han desarrollado varias tecnologías para disminuir esta limitación, incluyendo el despliegue de sensores portátiles, montados en tractores y vehículos aéreos, de reflectancia espectral y temperatura del dosel de los cultivos (Araus y Cairns, 2014; Zhao et al., 2019) (Figura 4).
 Figura 4. (Izquierda) Un tractor de alto despeje con múltiples sensores conectados se usa para fenotipar trigo en el programa del Dr. Jesse Poland en la Universidad Estatal de Kansas. Crédito de la foto: Pat Byrne. (Derecha) Un dron cuádruple (o UAV por sus siglas en inglés: vehículo aéreo no tripulado) captura datos de reflectancia espectral en una prueba de campo de trigo de la Universidad Estatal de Colorado. Crédito de la foto: Scott Reid.
Figura 4. (Izquierda) Un tractor de alto despeje con múltiples sensores conectados se usa para fenotipar trigo en el programa del Dr. Jesse Poland en la Universidad Estatal de Kansas. Crédito de la foto: Pat Byrne. (Derecha) Un dron cuádruple (o UAV por sus siglas en inglés: vehículo aéreo no tripulado) captura datos de reflectancia espectral en una prueba de campo de trigo de la Universidad Estatal de Colorado. Crédito de la foto: Scott Reid.
Otro hito en la historia del desarrollo de cultivos fue la comercialización de cultivos transgénicos (manipulados o modificados genéticamente) a mediados de la década de 1990. Estos cultivos contienen genes transferidos de otras especies mediante tecnología de ADN recombinante utilizando protocolos de laboratorio. Los cultivos transgénicos han tenido gran impacto en un número relativamente pequeño de cultivos, en particular maíz, soya, algodón, canola, alfalfa y remolacha azucarera. La mayoría de los cultivos transgénicos actualmente disponibles en el mercado han sido mejorados para conferirles tolerancia a herbicidas o resistencia a insectos, aunque también se han comercializado cultivos resistentes a virus, que no se oxidan y tolerantes a la sequía. La Academia Nacional de Ciencias, Ingeniería y Medicina de EE. UU. (2016) publicó un informe exhaustivo sobre el impacto de los cultivos transgénicos, incluyendo la seguridad para la salud y la seguridad ambiental. Con respecto a los parientes silvestres de cultivos, una preocupación ha sido la posible transferencia de transgenes a poblaciones silvestres. Este es un riesgo que variará dependiendo de cada cultivo y región geográfica, y debe ser considerado cuidadosamente antes de aprobar un cultivo transgénico para una nueva región.
La diversidad genética conservada en los bancos de germoplasma del mundo, en especial para los parientes silvestres de cultivos, es un recurso en gran parte sin explotar para el mejoramiento de cultivos (Byrne et al., 2018). Los desafíos para el mayor uso de parientes silvestres de cultivos en el desarrollo de cultivos fueron descritos por Dempewolf et al. (2017), basándose en consultas con expertos en 24 comunidades de cultivos. Estos desafíos incluyen datos fenotípicos y genotípicos insuficientes sobre las accesiones de parientes silvestres de cultivos; diferencias de ploidía y otras barreras de hibridación entre especies silvestres y germoplasma cultivado; la existencia de suficiente variación en el germoplasma cultivado de algunos cultivos; inferioridad de los parientes silvestres de cultivos en los rasgos deseados; arrastre por ligamiento y recursos humanos o financieros inadecuados para realizar la investigación y el desarrollo necesarios. La aplicación creativa de la tecnología y los cambios de política apropiados ayudarán a superar estos desafíos para desbloquear la diversidad de las colecciones de bancos de germoplasma para el mejoramiento de cultivos. Hübner y Kantar (2021) proporcionan guías para el muestreo en la naturaleza, la evaluación de las colecciones para determinar la variación favorable y la incorporación de beneficios genéticos en los programas de fitomejoramiento. Kashyap et al. (2022) proporciona ejemplos exitosos del uso de parientes silvestres en el mejoramiento de cultivos y discute estrategias para facilitar la integración de germoplasma silvestre a programas de mejoramiento.
4. Referencias
Araus JL, Cairns JE. 2014. Field high-throughput phenotyping: The new crop breeding frontier. Trends in Plant Science 19:52-61.
Byrne PF, Volk GM, Gardner C, Gore MA, Simon PW, Smith S. 2018. Sustaining the future of plant breeding: The critical role of the USDA-ARS National Plant Germplasm System. Crop Science 58:451-468.
Dempewolf H, Baute G, Anderson J, Kilian B, Smith C, Guarino L. 2017. Past and future use of wild relatives in crop breeding. Crop Science 57:1070-1082.
Gepts P. 2018. The domestication of our food crops. En: Chrispeels MJ, Gepts P, (editores). Plants, Genes & Agriculture: Sustainability through Biotechnology. Oxford University Press, Cary, North Carolina.
Harlan JR. 1992. Crops & Man, 2nd edition. American Society of Agronomy and Crop Science Society of America, Madison, WI.
Hübner S, Kantar MB. 2021. Tapping diversity from the wild: From sampling to implementation. Frontiers in Plant Science 12:626565.
IFPRI. 2002. Green Revolution: Curse or blessing? International Food Policy Research Institute, Washington, D.C.
Jain SM, Brar DS (editores). 2010. Molecular Techniques in Crop Improvement, 2nd edition. Springer, Dordrecht.
Kashyap A, Garg P, Tanwar K, Sharma J, Gupta NC, Ha PTT, Bhattacharya RC, Mason AS, Rao M. 2022. Strategies for utilization of crop wild relatives in plant breeding programs. Theoretical and Applied Genetics 135:4151-4167.
Murphy DJ. 2007. Plant Breeding and Biotechnology: Societal Context and the Future of Agriculture. Cambridge University Press, Cambridge, Reino Unido.
National Academies of Sciences, Engineering, and Medicine. 2016. Genetically Engineered Crops: Experiences and Prospects. The National Academies Press, Washington, D.C.
Schwanitz F. 1966. The Origin of Cultivated Plants. Harvard University Press, Cambridge, MA.
Sleper DA, Poehlman JM. 2006. Breeding Field Crops, 5th edition. Blackwell Publishing, Ames, IA.
Tanksley SD, McCouch SR. 1997. Seed banks and molecular maps: unlocking genetic potential from the wild. Science 277:1063-1066.
Voss-Fells KP, Cooper M, Hayes B. 2019. Accelerating crop genetic gains with genomic selection. Theoretical and Applied Genetics 132:669-686.
Zhao CJ, Zhang Y, Du JJ, Guo XY, Wen WL, Gu SH, Wang JL, Fan JC. 2019. Crop phenomics: Current status and perspectives. Frontiers in Plant Science 10:714.
5. Agradecimientos
Para citar este artículo: Byrne PF, Richards C, Volk GM. 2024. De especies silvestres a razas locales y cultivos. En: Volk GM, Byrne PF (Eds.) Parientes silvestres de cultivos y su uso en el fitomejoramiento. Colorado: Universidad Estatal de Colorado. [Escribe la fecha de acceso]. Disponible en: https://colostate.pressbooks.pub/cropwildrelativesspanish/chapter/de-especies-silvestres-a-razas-locales-y-cultivos/
Este módulo de capacitación fue posible, en parte, gracias al financiamiento del USDA-ARS, la Universidad Estatal de Colorado, IICA-PROCINORTE (procinorte.net) y la Agencia de Estados Unidos para el Desarrollo Internacional (USAID). Traducciones a español fueron apoyados por IICA-PROCINORTE.
Editores: Gayle Volk, Katheryn Chen
Traducido: Gabriela Sandoval Cancino, Andrea Purdy, Stephanie Villanueva
Camarógrafo: Mike May
Este proyecto fue financiado en parte por National Academy of Sciences (NAS) [la Academia Nacional de Ciencias] y USAID. Es de notar que las opiniones, los hallazgos, las conclusiones y las recomendaciones expresadas vienen de los autores y no reflejan necesariamente los puntos de vista de USAID ni de NAS.
